Peptide bond
This article is about the peptide link found within biological molecules, such as proteins. A similar article for synthetic molecules is being created. Although the atoms involved are the same, the terms amide link and peptide link cannot be used interchangeably.
A peptide bond (amide bond) is a covalent chemical bond formed between two molecules when the carboxyl group of one molecule reacts with the amino group of the other molecule, causing the release of a molecule of water (H2O), hence the process is a dehydration synthesis reaction (also known as a condensation reaction), and usually occurs between amino acids. The resulting C(O)NH bond is called a peptide bond, and the resulting molecule is an amide. The four-atom functional group -C(=O)NH- is called a peptide link. Polypeptides and proteins are chains of amino acids held together by peptide bonds, as is the backbone of PNA.
A peptide bond can be broken by amide hydrolysis (the adding of water). The peptide bonds in proteins are metastable, meaning that in the presence of water they will break spontaneously, releasing 2-4 kcal/mol [1] of free energy, but this process is extremely slow. In living organisms, the process is facilitated by enzymes. Living organisms also employ enzymes to form peptide bonds; this process requires free energy. The wavelength of absorbance for a peptide bond is 190-230 nm.[2]
Contents |
Resonance forms of the peptide group
The amide group has three resonance forms, which confer six important properties. First, it stabilizes the group by roughly 20 kcal/mol, making it less reactive than many similar groups (such as esters). The resonance suggests that the amide group has a partial double bond character, estimated at 40% under typical conditions. The peptide group is uncharged at all normal pH values, but its double-bonded resonance form gives it an unusually large dipole moment, roughly 3.5 Debye (0.8 electron-angstrom). These dipole moments can line up in certain secondary structures (such as the α-helix), producing a large net dipole.
The partial double bond character can be strengthened or weakened by modifications that favor one resonance form over another. For example, the double-bonded form is disfavored in hydrophobic environments, because of its charge. Conversely, donating a hydrogen bond to the amide oxygen or accepting a hydrogen bond from the amide nitrogen should favor the double-bonded form, because the hydrogen bond should be stronger to the charged form than to the uncharged, single-bonded form. By contrast, donating a hydrogen bond to an amide nitrogen in an X-Pro peptide bond should favor the single-bonded form; donating it to the double-bonded form would give the nitrogen five quasi-covalent bonds. (See Figure 3.) Similarly, a strongly electronegative substituent (such as fluorine) near the amide nitrogen favors the single-bonded form, by competing with the amide oxygen to "steal" an electron from the amide nitrogen (See Figure 4.)
Cis/trans isomers of the peptide group
The partial double bond renders the amide group planar, occurring in either the cis or trans isomers. In the unfolded state of proteins, the peptide groups are free to isomerize and adopt both isomers; however, in the folded state, only a single isomer is adopted at each position (with rare exceptions). The trans form is preferred overwhelmingly in most peptide bonds (roughly 1000:1 ratio in trans:cis populations). However, X-Pro peptide groups tend to have a roughly 3:1 ratio, presumably because the symmetry between the 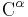 and
and 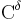 atoms of proline makes the cis and trans isomers nearly equal in energy (See figure, below).
atoms of proline makes the cis and trans isomers nearly equal in energy (See figure, below).
The dihedral angle associated with the peptide group (defined by the four atoms 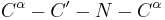 ) is denoted
) is denoted 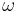 ;
; 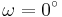 for the cis isomer and
for the cis isomer and 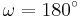 for the trans isomer. Amide groups can isomerize about the C-N bond between the cis and trans forms, albeit slowly (
for the trans isomer. Amide groups can isomerize about the C-N bond between the cis and trans forms, albeit slowly (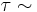 20 seconds at room temperature). The transition states
20 seconds at room temperature). The transition states 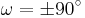 requires that the partial double bond be broken, so that the activation energy is roughly 20 kcal/mol (See Figure below). However, the activation energy can be lowered (and the isomerization catalyzed) by changes that favor the single-bonded form, such as placing the peptide group in a hydrophobic environment or donating a hydrogen bond to the nitrogen atom of an X-Pro peptide group. Both of these mechanisms for lowering the activation energy have been observed in peptidyl prolyl isomerases (PPIases), which are naturally occurring enzymes that catalyze the cis-trans isomerization of X-Pro peptide bonds.
requires that the partial double bond be broken, so that the activation energy is roughly 20 kcal/mol (See Figure below). However, the activation energy can be lowered (and the isomerization catalyzed) by changes that favor the single-bonded form, such as placing the peptide group in a hydrophobic environment or donating a hydrogen bond to the nitrogen atom of an X-Pro peptide group. Both of these mechanisms for lowering the activation energy have been observed in peptidyl prolyl isomerases (PPIases), which are naturally occurring enzymes that catalyze the cis-trans isomerization of X-Pro peptide bonds.
Conformational protein folding is usually much faster (typically 10-100 ms) than cis-trans isomerization (10-100 s). A nonnative isomer of some peptide groups can disrupt the conformational folding significantly, either slowing it or preventing it from even occurring until the native isomer is reached. However, not all peptide groups have the same effect on folding; nonnative isomers of other peptide groups may not affect folding at all.
Chemical reactions
Due to its resonance stabilization, the peptide bond is relatively unreactive under physiological conditions, even less than similar compounds such as esters. Nevertheless, peptide bonds can undergo chemical reactions, usually through an attack of an electronegative atom on the carbonyl carbon, breaking the carbonyl double bond and forming a tetrahedral intermediate. This is the pathway followed in proteolysis and, more generally, in N-O acyl exchange reactions such as those of inteins. When the functional group attacking the peptide bond is a thiol, hydroxyl or amine, the resulting molecule may be called a cyclol or, more specifically, a thiacyclol, an oxacyclol or an azacyclol, respectively.
References
- ^ Martin RB. (1998) "Free energies and equilibria of peptide bond hydrolysis and formation", Biopolymers, 45, 351–353.
- ^ Goldfarb AR et al. (1951) "The Ultraviolet Absorption Spectra of Proteins", J. Biological Chem., 193, 397-404.(http://www.jbc.org/content/193/1/397.long)
- Pauling L. (1960) The Nature of the Chemical Bond, 3rd. ed., Cornell University Press. ISBN 0-8014-0333-2
- Stein RL. (1993) "Mechanism of Enzymatic and Nonenzymatic Prolyl cis-trans Isomerization", Adv. Protein Chem., 44, 1-24.
- Schmid FX, Mayr LM, Mücke M and Schönbrunner ER. (1993) "Prolyl Isomerases: Role in Protein Folding", Adv. Protein Chem., 44, 25-66.
- Fischer G. (1994) "Peptidyl-Prolyl cis/trans Isomerases and Their Effectors", Angew. Chem. Int. Ed. Engl., 33, 1415-1436.
See also
|
||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||